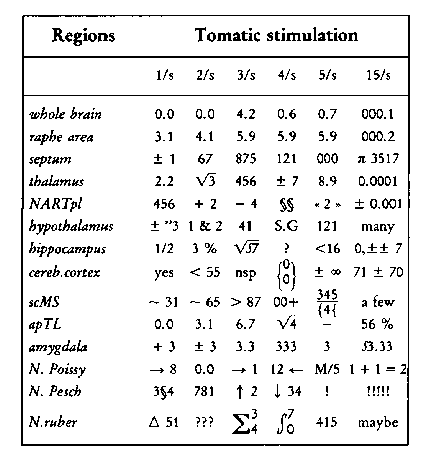
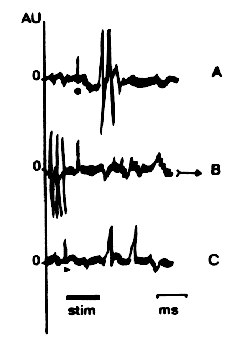
Alka-Seltzer, L. Untersuchungen über die
tomatostaltische Reflexe beim Walküre. Bayreuth Monatschr. f. exp.
Biol. 184, 34-43, 1815.
Attou, J. & Ratathou, F. Laminar configuration
of the thalamo-tomatic relay nuclei. Experimental study with Fink-Heimer-Gygax
methods. In: The Hyperthalamus, ed. by V. Cointreau and M. Brizard,
Cambridge, Oxford U.P., pp. 32-88, 1974.
Balalaïka, P. Deafness caused by tomato
injury. Observations on half a case. Acta. pathol. marignan. 1, 1-7,
1515.
Beulott, A., Rebeloth, B. & Dizdeudayre,
C.D. Brain designing. Châteauneuf-en-Thymerais, Institute of
advanced studies (vol. 17), 1974.
Bortsch, B. Saccular disturbances produced
by whistling (in russian). Fortschr. Hals-Nasen-Ohrenheilk. 3, 412-417,
1955.
Carpentier, H. & Fialip, L. Tomato calibres
& swallowing. Bull. diet. gastrom. Physiol. 3, 141-167, 1964.
Chachlik, I. Vocal performance and binoculars.
Covent Gard. J. 307, 1975-1080, 1959-1960.
Chou, O. & Lai, A. Tomatic inhibition in
the decerebrate baritone. Proc. koning. Akad. Wiss., Amst. 279, 33,
1927a.
Chou, O. & Lai, A. Note on the tomatic
inhibition in the singing gorilla. Acta laryngol. 8, 41-42,
1927b.
Chou, O. & Lai, A. Further comments on
inhibitory responses to tomato splitting in Soloists. Z. f. Haendel Wiss.
17, 75-80, 1927c.
Chou, O. & Lai, A. Faradic responses to
tomatic stimulation in the buzzling ouistiti. J. amer. metempsycb. Soc.
19, 100-120, 1928a.
Chou, O. & Lai, A. Charlotte's syndrome
is not a withdrawal reflex. A reply to Roux & Combaluzier. Folia pathol.
musicol. 7, 13-17 1928b.
Chou, O. & Lai, A. Tomatic excitation and
inhibition in awake Counteralts with discrete or massive brain lesions. Acta
chirurg. concertgebouw., Amst. 17, 23-30, 1929a.
Chou, O. & Lai, A. Musicali effetti del
tomatino jettatura durante il reprezentazione dell' opere di Verdi. In:
Festschrift am Arturo Toscanini, herausgegeb. vom A. Pick, I. Pick,
E. Kohl & E. Gramm., München, Thieme & Becker, pp. 145-172,
1929b.
Chou, O & Lai, A. Suprasegmental contribution
to the yelling reaction. Experiments with stimulation and destruction.
Ztschr. f. d. ges. Neur. u. Psychiat. 130, 631-677, 1930.
Colle, E., Etahl, E & Others, S. Leguminase
pathways in the brain. A new theory. J. Neurochem. Neurocytol. Enzymol.
1, 8-345, 1973.
Dendritt, A. & Haxon, B. Synaptic contacts
in the Lily Pons. Brain Res., 1975 (in the press).
Donen, S. & Kelly, G. Singing in the
brain. Los Angeles, M.G.M. Inc. Press. 1956.
Einstein, Z., Zweistein, D., Dreistein, V.,
Vierstein, F. & St. Pierre, E. Spatial integration in the temporal cortex.
Res. Proc. neurophysiol Fanatic Soc. 1, 45-52, 1974.
Else, K. & Vire, A. de. 45-years tomato
throwing on amateur Singers. New Records Ass. J. 27, 37-38, 1974.
Ford, G. Highways and pathways for motor control.
J. pyramid. Soc. 30, 30, 1930.
Giscard d'Estaing, V. Discours aux transporteurs
routiers de Rungis. C. r. Soc. fr. Tomatol. 422, 6, 1974.
Gorden, H.W. & Bogen, J.E. Hemispheric
lateralization of singing after intracarotid sodium amylobarbltone. J.
Neurol. Neurosurg. Psychiat. 37, 727-738, 1974.
Harvar, D. & Mercy, B.C.P. Reward and
punishment in Olympic throwers. Hammersmith J. 134, 12-15, 1973.
Heinz, D. Biological effects of ketchup splatching.
J. Food Cosmet. Ind. 72, 42-62, 1952.
High, A.B.C.D. & Low, E.F.G.H. Cerebellar
aphonia and the Callas syndrome. Brain 91, 23-1, 1968.
Hubel, D.H. & Wiesel, T.N. Receptive &
tomato fields in the zona incerta. Experientia 25, 2, 1970.
Hun, O. & Deu, I. Tonic, diatonic, &
catatonic stage-distress syndromes. Basel, Karger, 1960.
Jeanpace, L. & Desmeyeurs, P. Recherches
histologiques sur les noyaux de Pesch & de Poissy. Dijon med. 5,
1-73, 1932.
Karybb, H. & Szyla, H. Of birds and men:
calling strategies and humming responses. Biol. Gaz. Elec. 73, 19-73,
1973.
Kuffler, S.W. Papezian control of aggressive
borborygms in Julliard drop-out. J. physiol. Physiol. 2, 21 -42,
1969.
Lai, A. & Chou, O. Dix-sept recettes faciles
au chou et a l'ail. I. Avec des tomates. J. Ass. philharmon. Vet. lang.
fr. 3, 1-99, 1931a.
Lai, A. & Chou, O. Dix-sept recettes faciles
au chou et a l'ail. II. Avec d'autres tomates. J. Ass. philharmon. Vet.
lang. fr. 3, 100-1, 1931b.
Loewenstein, W.R., Lowenfeld, I., Lövencraft,
N., Løwoenshrift, Q. & Leuwen, X. Tomatic neuralgia. J. Neurosurg.
Psychiat. Neurol. 340, 34-89, 1930. |
Mace, I. & Doyne, J. Sur les différents
types de réactions tomateuses chez la Cantatrice. Gaz. méd.
franco-rus. 6, 6-11, 1912.
Malosol, T. Utricular responses during tomato
conditioning. Bull. med. Aunis & Saintonge 43, 6-11, 1956.
Maotz, E. & Toung, I. Tomatic innervation
of the nucleus ruber. Proc. Opossum Soc. 70, 717-727, 1973.
Marks, C.N.R.S. & Spencer, D.G.R.S.T. About
the frightening reactions that accompanied first performances of Il
trovatore at the Metropolitan. Amer. J. music. Deficiency 7, 3-6,
1899.
Mason, H.W. & Rangoun, S.W. Paratrigeminaloid
musicalgia. In: 3rd Conference on the Rimsky-Korsakoff syndrome, ed.
by T. Thanos & P. Roxidase, Springfield, Ill., C. C. Thomate, pp. 31-57,
1960.
McCulloch, W.S., Pitts, W.H. & Levin, R.D.Jr.
What's the frog stomach tells to the frog's audience. Proc. Leap &
Frog Ass. 64, 643-1201, 1964.
McHulott, E., Mac Haskett, E. & Massinture,
.T.C. Fate of exogenous (14C) scotch, (235U) bloodymary and other tritiated
compounds injected in laryngeal and pharyngeal pathways. Clin. Bull. B.P.R.
Soc. 89, 35-78, 1975.
Others, S., Colle, E. & Etahl, E. The enzymase
enigma revisited. Am. J. Allegrol. 43, 234-567, 1974.
Otis, J. & Pifre, K. Gasping in the ascending
pathways. In: Hommage à Henri Eiffel, ed. by D. Haux &
D. Bas, Paris. C.N.R.S., pp. 347-950, 1964.
Payre, L. & Tairnelle, E. Sur le sursaut
tomateux du Baryton léger. C.R. Assoc. Conc. Lam. 45, 6-7,
1916.
Pericoloso, O. & Sporgersi, I. Sull'effetti
tomestetiche e corticali della stimolazione di leguminose nella Diva. Arch.
physiol. Schola Cantor. 37, 1805-1972, 1973.
Pesch, U. Experimentelle Beitrage uber anterior
reticularis Kerne beim Minnesanger. Von Bulow's Arch. f. d. ges. Musikol.
1, 1-658, 1876.
Poissy, N. de. Atrophie congénitale
des Noyaux de Pesch. Bibl. clin. Homeoprat. Lugdun. 65, 22-31,
1880.
Pompeiano, O., Vesuviana, A., Strombolino,
H. & Lipari, G. Volcaniche effetti della formazione reticolare nella
funiculi funicula. C.r. Ass. ital. Amat. Bel Cant. 37, 5-32,
1971.
Remmers, J.E. & Gauthier, H. Neural and
mechanical mechanisms of feline purring. Respir. Physiol. 16, 351-361,
1972.
Roux, C.F. & Combaluzier, H.U. Le syndrome
de Charlotte. Weimar Ztschr. musikol. Pomol. 7, 1-14, 1932.
Sinon, E., Evero, I & Ben Trovato, A.
Psychopathological description of La Furia di Caruso (in italian).
Folia clin. oto-rhinolaryngol., Foum Tataouine 6, 362-363, 1948. (Quoted
by Hun & Deu, 1960).
Sornette, U. & Billevayzé, H. Les
stomatites tomateuses. Arch. municip. Météorol. lyr.
Déontol. music. 264, 14-18, 1925.
Strogonoff, H., III. Pineal activation and
the yelling reaction. Show Busin. med. Gaz. 3, 273-308, 1960.
Sturm, U. & Drang, F. Musikalische
Katastrophe. Berlin, W. de Gruyter, 1973.
Szentagothai, J. The substantia nigra as a
striatal machine. Bull. Ass. niger. Neurophysiol. clin. exp., Niamey 23,
25-40, 1972.
Tarama, K. Acid-base balance. pHD Thesis,
San Francisco, 1957.
Tebaldi, R. La Callas revisited. Metropolitan
J. endocrin. Therap. 6, 37-73, 1953.
Timeo, W., Danaos, I. & Dona-Ferentes,
H.E.W. Brain cutting and cooking. Arch. metaphys. endogen. Gastrom. 56,
98-105, 1971.
Unsofort, H & Tchetera, K.G.B. Shout and
Yell. Yale J. Med. 9, 9-19, 1973.
Van der Deder, J. Von. The yelling pathway.
San Diego J. exp. Teratol. 50, Suppl. 24, 1 -28, 1950.
Vincent, J., Milâne, J., Danzunpré,
J.J. & Sanvaing-Danlhotte, J.J.J. Le réflexe hydro-musical. Gaz.
med. Faidh. Chalign. & d.s. Fil., 1976 (in the press).
Von Aitick, A. Ueber geminal-niebelungenischen
Schmerz. Ztschr. exp. pathol. Tomatol. 4, 4a-64P, 1940.
Wait, H. & See, C. Balistic requirements
in tomato throwing and splatching. Nasa Rept. N°68/67S/002/F4,
1-472, 1972.
Wimbledon, A.F.G.H. On the Statistical matching
of neuronal and other data. J. dynam. Stat. 5, 1-28, 1974.
Zakouski, B.G.H. Investigations d'avant-garde
sur les voies fluviales artificielles à moitié rondes dans
le hall d'entrée (traduit du russe). Exp. J. sechenov. Pflugerol.
3, 17-34, 1954.
Zeeg, O & Puss, I.K. On the fanatic
demonstrations of music lovers. J. behav. developm. Psychobiol. 31,
1-13, 1931.
Zubrowska, A. Oculo-tomatic dyskinesia. A
preliminary report. J. neuro-neurol. Neurol. 1, 107, 1958.
Zyszytrakyczywsz-Sekrâwszkiwcz, I. The
Monte Carlo theorem as a use in locating brain and other sites. J. math.
Vivisec. 27, 134-143, 1974. |